|
|
|
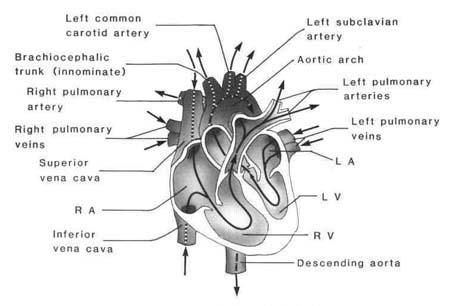
Figure 1.1 Blood flow through the heart |
| Contents | Previous | Next |
The heart is a four-chambered pump, lying in the mediastinum in the thoracic cavity between the lungs. Functionally, the heart is divided into a right and a left pump. The upper chambers, called atria, act as collectors for the lower chambers, called ventricles. Ventricles produce the heart's pumping action. The right heart receives blood from the systemic veins and propels this venous blood to the lungs for oxygen supply and carbon dioxide removal. The left heart pumps oxygenated blood received from the pulmonic veins into systemic circulation.
The pericardium is a tough fibrous sac that encloses the heart and holds it in place by attaching it to the sternum anteriorly, to the vertebral column posteriorly, and to the diaphragm inferiorly. Two layers, a parietal or outer layer and a visceral or inner layer, make up the pericardial sac. A small amount of fluid (approximately 8-10 ml) between the layers acts as a lubricant, allowing the heart to beat without friction.
The base of the heart faces up and to the right, and the tip, or apex, faces down and to the left. Identification of the anatomic structures and surfaces that make up the areas of the heart is important to the understanding of abnormalities. Because the heart is rotated slightly to the left, when viewed in the frontal plane, the right atrium and right ventricle lie anteriorly and in front of the left atrium and left ventricle. The left ventricle comprises the largest section of the inferior (diaphragmatic), left lateral, and apical surface of the heart. The anterior and right lateral surface comprises primarily the right ventricle with a small section of right atrium and left ventricle. The apex, called the point of maximal intensity (PMI), is formed by this small section of left ventricle. The PMI is the point of maximal outward movement of the precor-dium during systole. The posterior surface is composed of small sections of the right atrium and right ventricle and the left atrium (Fig. 1.1).
|
|
|
Figure 1.1 Blood flow through the heart |
The right atrium is a low-pressure, thin-walled chamber, functioning as a reservoir and conduit for systemic venous blood. Blood enters the right atrium via the superior vena cava, the inferior vena cava, and the coronary sinus, which drains venous blood from the heart itself. Three layers make up the walls of the atria: (1) epicardium, a thin outer layer adjacent to the pericardium, continuous with the outer layer of the ventricles; (2) myocardium, the muscular middle layer, discontinuous with that of the ventricles; and (3) endocardium, the thin smooth endothelial layer that makes up the inner layer. This continuous layer covers the inner surface of the two atria, the valves, the ventricles, and the great vessels. The low pressure in the atria is accounted for by the thinner muscular layer of the atria and thicker wall muscle in the ventricles. Blood flows from the right atrium into the right ventricle through the tricuspid valve. Normally, this flow accommodates about 80% of the venous blood. The atrial contribution to cardial output, called the atrial kick, adds approximately 20% more to ventricular filling. No true valves separate the vena cava and right atrium, so any pressure changes in the right atrium are reflected back into the venous circulation.5
Three layers also make up the ventricular walls: the epicardium, the myocardium, and the endocardium. The workload of the right ventricle is far lighter than that of the left. The right ventricle pumps against the low pressure pulmonic circulation, forcing blood into the pulmonary artery. The right ventricular myocardial layer is one-third the thickness of the left ventricular wall myocardium.
The left atrium is a low-pressure chamber slightly smaller than the right atrium. Oxygenated blood is received from the lungs into this chamber via four pulmonary veins. Valves do not exist between the pulmonary veins and left atrium, so changes in left heart pressures are reflected by the pulmonary circulation.
Blood from the left atrium moves into the left ventricle through the mitral valve. The greatest percentage of the heart's muscular tissue is found in the left ventricle. The inner aspects of this wall are crisscrossed with raised heavy muscle bundles called trabecular carnae cordis. The thickness of the wall permits the left ventricle to generate enough pressure to pump blood throughout systemic circulation.
The intra-atrial and intraventricular septum anatomically divide the right and left heart.5, 58 The membranous atrial septum divides the right and left atrium, preventing communication between the two. Within the septum is the fossa ovalis, the original fetal communication called the foramen ovale. The atria and ventricles operate independently in sequence through the continuation of the membranous septum. The atrioventricular groove creates the external separation. The right and left heart are further separated by the septum. A thin wall separates the atria, and a much thicker muscular wall separates the ventricles. The septum also contains the branches of the conduction tissues of the heart.
The inner surface of the ventricles houses the muscle bundles called trabeculae carnae, which contain two groups of papillary muscles in the left ventricle and three groups in the right ventricle. The chordae tendinae of the atrioventricular valves originate here.
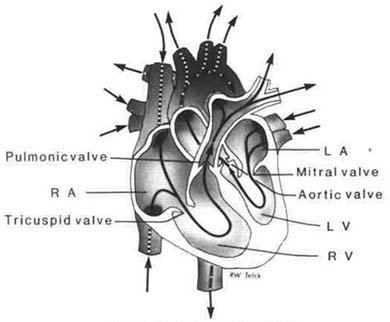
Unidirectional blood flow is controlled through the chambers of the heart by four valves. These valves are thin cusps, or leaflets, of fibrous tissue, continuous with the endocardium. The tricuspid valve between the right atrium and right ventricle consists of three leaflets. These leaflets are thinner than those found in the mitral valve of the left heart. The mitral valve separates the left atrium from the left ventricle and is composed of two leaflets. The valves are connected by the chordae tendinae, thin fibrous tissue strands, to the papillary muscles, which are myocardial projections from the ventricular wall. The chordae tendinae are attached to the cusps and to the papillary muscle to anchor the valves during ventricular contraction, completing closure and preventing eversion into the atria when blood is propelled forward during systole (contraction).
|
|
|
Figure 1.2 Valves of the heart |
The aortic and pulmonic valves are referred to as semilunar. They are similar to each other, with three symmetrical cuplike cusps. These valves do not have chordae tendinae or papillary muscles; they are instead secured to a fibrous ring. The aortic valve is found between the aorta and the left ventricle, with attachment to the walls of the base of the aorta. The pulmonic valve is found between the pulmonary artery and the right ventricle, attached to the pulmonary artery wall. These valves prevent backward flow of blood from the aorta and the pulmonary artery into the ventricles during ventricular diastole (relaxation).
During ventricular systole, the increased pressure in the ventricles forces the mitral and tricuspid valves to close. As pressure increases rapidly in the ventricles, it will exceed the pressure in the pulmonary artery and the aorta, forcing the pulmonic and aortic valves to open. The blood in the ventricles is thus expelled into systemic and pulmonary circulation. The pressure in the ventricles is then reduced, closing the semilunar valves and preventing backward flow. The period between the closing and opening of these valves is called diastole. Filling of the ventricles occurs at this time as the mitral and tricuspid valves open.
The inflow channels of the heart, the vena cava and the pulmonary veins, are not separated from the atria by valves. Blood flow into the heart is continuous. Distention of the atria, right or left, results in backward flow into systemic or pulmonary venous circulation, respectively.
The cardiac cycle describes the contraction and relaxation of the heart. Diastole is the filling period in which the heart receives blood. Systole is the ejection period.2, 5, 14 During diastole, the volume in the ventricles is normally increased to about 120 ml. This is referred to as the end diastolic volume. Approximately 50 ml remain in the ventricles at the end of systole. The difference between the diastolic volume (end diastolic volume) and the remaining volume following systole (end systolic volume) is the stroke volume. Normal stroke volume is about 70 ml. Cardiac output is determined by multiplying stroke volume times heart rate (SV x HR = CO) (see Chapter 4). Two factors that influence stroke volume, other than contractility, are preload and afterload. Preload is alternately described as filling volume, ventricular filling pressure,9 or the degree of stretch of myocardial muscle just prior to contraction.35 Preload relates to factors that occur before the electrical excitation and contraction. Decreased venous return to the right heart means a decrease in preload.
The force against which the ventricle must pump after the contraction has begun is called afterload.10 It refers to the resistance that is normally met in the aorta, the pulmonic valve, and the systemic vasculature. Increased viscosity of blood, added preload, hypertension, aortic stenosis, and idiopathic hypertrophic subaortic stenosis (IHSS) contribute to afterload by increasing the pressure against which the ventricle must pump.
Termed the great vessels, the major arteries of the heart are the aorta and the pulmonary artery. Separated from the left ventricle by the aortic valve, the aorta carries oxygenated blood to all of the body's tissues. The aorta branches numerous times to nourish systemic tissues. The pulmonary artery, from the right ventricle to the lungs, branches into smaller sections, finally becoming the pulmonary capillary bed. The exchange of oxygen and carbon dioxide occurs at this level. The pulmonic valve is the entry point from right ventricle to pulmonary artery.
Three layers make up the aorta and the pulmonary arteries: (1) an endothelial layer, or tunica intima; (2) a muscular layer called the tunica media; and (3) an adventitial, or outer layer, called the tunica externa.
The coronary circulation, branches of small arteries carrying oxygen and nutrition to the myocardium, supplies the heart's metabolic requirements. Damage to any one of these arteries can seriously impair the function of the heart muscle and the conduction system.
The coronary arteries are the heart's blood supply. These arteries arise from the sinuses of Valsalva, just above the aortic valve. The major coronary arteries are the right coronary artery and the left main coronary artery. The right coronary sinus gives rise to the right coronary artery (RCA), and the left coronary sinus gives rise to the left coronary artery (LCA).
The right coronary artery is usually a single vessel. When a second artery arises from the right coronary sinus, it is called the conus artery. When a single artery arises on the right, the first branch of the right coronary artery is then termed the conus artery.58
The left main coronary artery arises as a single artery before it divides, or bifurcates, into its two major branches, the left anterior descending and the left circumflex. Diagonal arteries may also branch off at the bifurcation area of the left coronary artery. These diagonal arteries may number as many as four.
The coronary arteries extend over the epicardial surface of the heart. Branches occur many times as the arteries penetrate the myocardium. These collateral arteries join the main coronaries via anastomatic channels. Blood supply to the epicardial myocardium is greater than that to the inner, or endocardial, layer because of the lie of the main coronary arteries. The lesser blood supply results in greater vulnerability to heart damage at the endocardial level. The coronary arteries circle the heart via external grooves. The right coronary artery moves in the atrioventricular groove and turns downward in the posterior surface to the posterior interventricular groove. The point at which the grooves (interventricular and atrioventricular) join the posterior heart surface is called the crux of the heart. The atrioventricular (AV)28, 58 node is found at this point and gains its blood supply from the vessel that crosses this point. Left dominance or right dominance is determined by this vessel crossing. The term dominant is used to designate the coronary artery that becomes the posterior descending artery.
In 55% of the population, the sinoatrial (SA) node, in the right atrium, is supplied by the right coronary artery. For perhaps 45% of the population, the circumflex artery is the major supplier of blood to the SA node. In a small part of the population (10%), the posterior descending artery is formed by the left circumflex artery rather than by the right coronary artery.
The left anterior descending (LAD) artery passes through the anterior interventricular groove, supplying blood to portions of the left and right ventricular myocardium, the anterior papillary muscle of the left ventricle, the anterior interventricular septum, and the apex of the heart. The circumflex artery, at a sharp angle coming off the left main coronary artery, moves to the left and posteriorly in the atrioventricular groove, traveling to the posteriolateral surface of the left ventricle. Its branches supply blood to the left ventricular lateral wall and parts of the left atrium.
|
|
|
Figure 1.3 Coronary Circulation. |
The right coronary artery, lying in the right atrioventricular groove, moves to the back of the heart, where it usually becomes the posterior descending artery. This artery supplies the right atrium, the right ventricle, and a part of the inferior and posterior surfaces of the left ventricle. It supplies blood to the His bundle and to the AV node in about 90% of the population.
The coronary arteries divide into smaller branches to move into the myocardium. This branching forms a network of capillaries that supplies the myocardium at the cellular level.
The large veins of the heart lie in close proximity to the arteries. The largest veins lie parallel to the circumflex artery flowing into the great cardiac vein, which becomes the coronary sinus. Venous drainage of the myocardium is provided by these veins. The majority of myocardial venous blood is then emptied into the right atrium through the coronary sinus os-tium, proximal to the orifice to the inferior vena cava. Drainage from the left heart travels to the coronary sinus and its branches. Venous drainage from the right atrial and right ventricular myocardium is via smaller veins, the the besian veins, that empty directly into the right atrium.
The autonomic nervous system (ANS) provides an external influence on activity of the conduction system, contractile properties of the heart, and heart rate. The parasympathetic nervous system (PNS) and the sympathetic nervous system (SNS) make up the ANS. Contractility and heart rate are adjusted by the ANS to changing body demands. The inherent pace-maker system of the heart is also influenced—enhanced or restrained. Automaticity of ectopic pacemakers may also be altered.
Sympathetic nervous system fibers are found in the atrial wall, the ventricles, and the SA and AV nodes. Stimulation results in the release of norepinephrine.14 This action stimulates the rate of depolarization and impulse transmission through the conduction tissues. When sympathetic stimulation is increased, heart rate and contractility increase. Normally, the sinus node is the recipient of increased stimulation. Fibers from the sympathetic nervous system occur in the SA and AV nodes as well as in the walls of atria and ventricles. Stimulation of the fibers increases the rate of depolarization and impulse transmission through the conduction system. Sympathetic stimulation accelerates heart rate, firing of the sinus node, conduction through the AV node, and the force of cardiac contraction. Sympathetic stimulation releases norepinephrine and epinephrine; this is referred to as adrenergic response.
Activity of the parasympathetic nervous system (PNS) is through acetylcholine, a chemical transmitter, released from the vagal fibers. The vagus nerve carries impulses from the PNS to the heart, mainly to the SA node, the AV node, and the atrial muscle. Parasympathetic stimulation slows the heart rate, the rate of impulse formation in the SA node, and conduction through the AV node. Excess stimulation of the vagus nerve can hinder the SA pacemaker and block impulse transmission through the AV node. Slowing of the heart rate results from reduced diastolic depolarization in the conduction system. Parasympathetic stimulation with the release of acetylcholine is referred to as a cholinergic response.
Regulation of the cardiovascular system by the autonomic nervous system requires several components: (1) sensors; (2) afferent pathways; (3) an integration center; (4) efferent pathways; and (5) receptors.
Sensors are the pressure sensitive structures called baroreceptors and chemoreceptors.5, 14, 58 The baroreceptors found in the carotid sinus and the aortic arch respond to decreased systolic blood pressure with a reflex sympathetic response of increased contractility, heart rate, and vasoconstriction. Increased pressure stimulates the receptors, and a reflex vagal response results in decreased heart rate; inhibited cardiac activity occurs. The medulla oblongata is the major chemoreceptor, but special receptor sites are found in the carotid body and aortic arch. Stimulation of these sensors occurs as a result of chemical changes in the blood, particularly arterial oxygen concentration, carbon dioxide, or pH (hydrogen ion concentration). Decreased p02or pH causes a sympathetic response, resulting in increased heart rate, contractility, and vasoconstriction. Decreased pCO2 and increased pH reduces vasoconstrictor effects, leading to a general vasodilatory effect.
The neural impulses are carried from the receptors to the brain via afferent pathways in the vagus and glossopharyngeal nerves. The lower pons and upper medulla act as integration centers for cardioregulatory effects. The integrated center receives messages from the baro- and chemoreceptors and sends messages to the cardiovascular system via the parasympathetic and sympathetic nerve fibers. For the parasympathetic fibers, the vagus nerves are the efferent pathways from the cardioregulatory center to the heart. The sympathetic fibers use the cardiac nerves. The receptors are found in the myocardium, the conduction system, and the smooth muscle of blood vessels.
In the cardiovascular system, sympathetic (adrenergic) receptor sites are divided into alpha and beta adrenergic receptors. Increased alpha adrenergic activity causes vasoconstriction of blood vessels in splanchnic, skin, and cerebral circulation. Beta receptors are further divided into beta1 (β1) and beta2 (β2). Beta1 receptors are found in the heart, β2 receptors in the lungs. Stimulation results in an increased heart rate, contractility, and increased activity of the SA node, which, in turn, increases the heart's irritability. Up to a point, increased heart rate and contractility, with adequate venous return, will increase cardiac output and elevate blood pressure. Stimulation of β2, receptors of blood vessels results in vasodilatation of these blood vessels in smooth muscle cells.
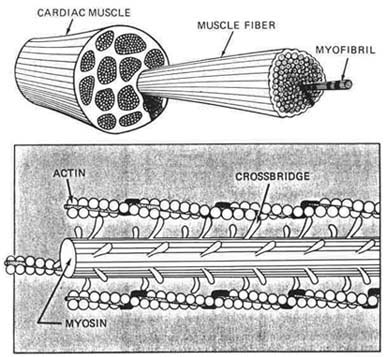
There are differences in structure and function between skeletal muscle, cells and cardiac muscle cells. Cardiac muscle cells have one central nucleus; skeletal cells have peripheral nuclei. In cardiac muscle, the cells are close together, so impulses pass quickly from one cell to the next. A cardiac muscle comprises many muscle fibers. The muscle fiber contains tiny tube like structures called myofibrils.
The myofibril is the contractile unit of a myocardial muscle cell, the basic unit of cardiac muscle fiber. The sarcomere is the basic contractile unit of a myofibril. Myofibrils are composed of many sarcomeres (Fig. 1.4). Each myofibril is composed of two kinds of protein filaments called actin and myocin, which lie side by side and are connected by cross bridges. Action occurs through movement of the actin on myocin in the sarcomere unit. The actin and myocin filaments slide together and overlap for muscle contraction and move apart for relaxation. The thin filament, actin, contains two proteins, tropomyocin and troponin, which inhibit activity of the filaments.34
|
|
|
Figure 1.4 Contracting elements. |
Electrical excitation of the myocardial cells creates the muscular contraction. This occurs as a result of calcium diffusion through the sarcoplasm of the cell to the sarcomere. Calcium binds with troponin, one of the two actin proteins, inactivating the inhibitory action of the two proteins and causing the actin and myocin myofilaments to interact. This interaction, the sliding filament theory, is the overlap activity that produces muscular contraction. Relaxation occurs when the calcium is taken up by the sarcoplasmic reticulum, causing actin and myocin to move apart. The energy for this activity comes from adenosine triphosphate (ATP), which is split by an adenosine triphosphatase (ATPase) site on the myocin filament when it interacts with actin. The force of the contraction is dependent on this interaction between sarcomere myofilaments.
Cardiac muscle cells lie quite close to one another. A tight junction between cells occurs via intercalated disks, allowing for rapid conduction of impulses between the cells. This cellular structure permits the entire myocardial unit to respond when one cell is stimulated to threshold level.
Mitochondria, small rod-shaped structures, are literally the power plants of a cell. They are the sites where energy from nutrients is transformed. The mitochondria are located close to the myofibrils in muscle cells. The large numbers of mitochondria in cardiac muscle is determined by the high activity level and energy needs of cardiac cells. Cardiac muscle requires a great deal of ATP to function properly. ATP is produced in the myocardial mitochondria, using fatty acids, lactate, and glucose. The production of ATP varies with the workload of pumping blood into the arteries. Cardiac work may be expressed in terms of oxygen consumption. Factors influencing oxygen consumption include: (1) contractile state; (2) myocardial muscle mass; (3) heart rate; and (4) amount of tension generated within the myocardium. The coronary arteries deliver the required oxygen. When a situation involving stress arises, the oxygen demand rises in response to increased heart rate and increased contractility produced by the catecholamines.
The electrical activity developed in a cardiac muscle cell occurs as the result of movement of ions across the cell membrane and is termed the action potential.5, 28 Because a cell membrane is semipermeable, ions of opposite charge become aligned on either side of the cell membrane. The charge is negative in a resting state. The charged ions flow through ion selective channels found along the membrane, referred to as ion fast or ion slow channels. The specific ions related to cardiac electrophysiology are sodium (Na+), potassium (K+), calcium (Ca++), and chloride (C1-). Normally, the intracellular fluid contains a concentration of 150 mEq per liter of K+, and the extracellular fluid contains about 3-5 mEq per liter. Na+ concentration is extracellular, normally the reverse of K+. Action begins with a stimulus to the cell membrane that becomes permeable to Na+. A rapid influx of Na+ occurs, changing the negative charge of the polarized cell to a positive charge. This change, called depolarization, is Phase 0 of the action potential. It is believed that a fast channel opens, permitting the rapid influx of Na+. Ca++ ions move at the same time as the Na+ ions, with one Ca++ ion moving in as two Na+ ions move. A small influx of Cl- accompanies the Na+ influx. In an attempt to restore equilibrium, K+ leaves the cell as the fast Na+ channel begins to close, reducing the Na+ influx. Phase 1 is a period of rapid depolarization. The plateau, or Phase 2, is the more gradual depolarization phase. The slow channels seem to carry the activity with small amounts of Na+movement. Phases 3 and 4 involve restoring ion balance via a pumping mechanism that moves excess Na+ ions out of the cell while the K+ ions return to original levels. This action is referred to as the sodium-potassium pump.
Phase 4 is the resting phase, or repolarization state, with intracellular ion concentration maintained. The cell is again ready to accept an electrical stimulus. The action potential in the four phases described occurs in the myocardial cells of the atria, ventricles, and the His-Purkinje system. This is the fast channel. Entry into the cell of Ca+ + ions is via pathways designated as slow channels. This is another type of action potential that occurs in the cells of the AV and SA nodes. This influx occurs over a longer period and couples Ca+ + ions with entering Na+ ions. Ca+ + is necessary to create a contraction. When there is insufficient intracellular Ca+ +, cardiac contractions can be weakened. (See Section Five, Chapter 14 for further discussion of Ca++ and action potential.)
Heart muscle possesses several properties that help to ensure synchronized, rhythmic excitation and contraction. Specialized conduction pathways lie in the myocardium, exhibiting the properties of:
|
|
|
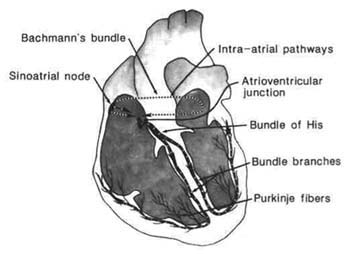
Figure 1.5 Conduction system. |
These properties create the difference in heart muscle, allowing it to beat independently of the nervous system. Generating its own electrical impulses (action potentials) rhythmically and spontaneously, these impulses are then carried through the conduction system to create muscular contraction of the myocardium. Specialized conduction tissues initiate normal contraction of the heart. The structures included in the process are the sinoatrial node (SA), the atrial internodal tracts, the atrioventricular junction (AV), the His bundle, the right and left bundle branches, and the Purkinje fibers. Cardiac contractions normally originate in the specialized tissue of the SA node, where there is a leakage of Na+ into the cell. Because this is not countered by an electrically equal positive ion moving out of the cell, initiation of the action potential, or diastolic depolarization, occurs.
Parasympathetic influences via the vagus nerve can alter SA node automaticity, slowing the rate of depolarization and decreasing the automaticity. Sympathetic influences also alter SA node automaticity, increasing the rate of the depolarization.28, 58
The SA node is referred to as the normal pacemaker of the heart. It is found in the posterior wall of the right atrium near the entry area of the superior vena cava. This specialized tissue acts as the heart's pacemaker because its intrinsic firing rate is faster than other parts of the conduction system. The impulse travels from the SA node through the atrium by specialized atrial conduction pathways, the atrial internodal tracts. Bachmann's bundle, one interatrial tract, carries the impulse to the left atrium. Other internodal tracts, the anterior, middle, and posterior, carry the impulse to the AV node. The AV node is
positioned in the right atrium near the opening of the coronary sinus at the top of the interventricular septum. Junctional fibers in the AV node connect with the His bundle. This node has two functions: delay of the cardiac impulse to allow the ventricles to fill and control of the atrial impulses that are conducted to the ventricles. This controlling influence protects the ventricles when atrial rates exceed normal limits.
Atrial rates occasionally can reach more than 400 beats per minute (bpm). Accelerated atrial rates produce ventricular filling problems. When tachycardia is present, the time spent in diastole is reduced, with less time for ventricular filling prior to systole. Another consideration is the ratio of atrial to ventricular beats. For example, atrial flutter may have a 4:1 conduction ratio. This can produce a ventricular rate of 75 bpm. Several problems can arise when there is a fast atrial rate: (1) loss of atrioventricular synchrony with the atria contracting against closed AV valves;(2) loss of the atrial kick; or (3) incomplete atrial emptying, which can lead to blood stagnation with clot or emboli formation.
From the AV node, the impulse is carried by specialized conduction fibers into a shaft called the common bundle, or His bundle, a thick bundle of fibers extending down the right side of the interventricular septum. Kent bundles, accessory tracts outside the AV node and His bundle, have also been identified. The His bundle separates as it reaches the interventricular septum into right and left bundle branches, which descend on opposite sides of the septum. The left bundle branch bifurcates into three fascicles: a thin anterior division called the left anterior fascicle, a thick posterior division called the left posterior fascicle, and a septal fascicle. The left bundle descends via the left interventricular septum. The anterior fascicle travels to the anterior and lateral walls of the left ventricle. The posterior fascicle goes to the lateral and posterior walls and to the papillary muscle. The septal fascicle moves to the interventricular septum and the apex of the left ventricle.
The right bundle branch is a single set of fibers descending superficially in the endocardium of the right ventricular septum. The walls of the right ventricle are penetrated by small branches of the right bundle. The bundle branches continue through subendocardial tissues toward the papillary muscles, terminating in a network of fibers. The Purkinje fibers provide a continuation of the bundle branches. Made up of large fibers, the Purkinje system allows for rapid conduction and simultaneous excitation of the inner surface of both ventricles.
Normally, an impulse moves through the conduction system in an orderly fashion. Starting at the SA node, the impulse is carried via the atrial pathway to the AV node. His bundle, bundle branches, and Purkinje fibers. Impulses usually begin in the SA pacemaker. In the event that the SA node fails as an impulse generator, other tissues assume the pacemaker role. The AV junction may take over with an intrinsic rate of 40-60 bpm, or a ventricular site in the Purkinje fibers may become the pacemaker with a 20-40 bpm rate. These auxiliary pacemakers can prevent asystole or cardiac standstill when the SA node is dysfunctional.
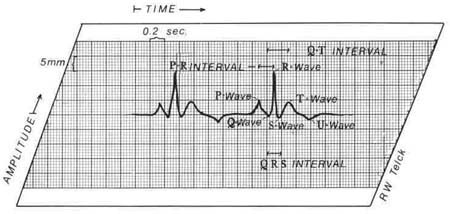
The electrical activity of the heart (Fig. 1.6) can be detected on the body surface and displayed as waveforms. This recording is called an electrocardiogram (ECG). The paper used to record an ECG is standardized, so when lines, areas, or boxes are measured, everyone is describing the same feature. The horizontal lines of the ECG paper represent time. Descriptors refer to large and small boxes. A small box represents .04 seconds, and a large box represents .20 seconds. Voltage or amplitude is measured by the vertical lines. One small box is equal to 1 mm, which equals . 1 millivolt (mV), and one large box is equal to 5 mm. The reflections are called waves or complexes. They have measurable intervals. An isoelectric line represent sents no electric forces in motion. Positive waves are those above the line, and negative waves are those below the line. Waves that are both positive and negative are called biphasic. Monitoring from different leads, or positions, creates the positive or negative waveform.
|
|
|
Figure 1.6 ECG monitor strip with each waveform shown. |
This wave represents atrial muscle depolarization. It is normally .11 seconds or less in duration. Normal amplitude is 2.5 mm or less.
The first negative deflection after the P wave is the Q wave, less than .03 seconds; the first positive deflection is the R wave. The S wave appears as the first negative deflection after the R wave. The QRS complex represents ventricular muscle depolarization from the beginning of the Q wave to the end of the S wave, normally .06-.11 seconds. When this complex is all negative, it is referred to as a QS complex. All of the waveforms, Q, R, S are not always present in a QRS complex. The waveform pattern will depend on the lead in use and the position of the heart.
The T wave represents ventricular muscle repolarization, the resting phase in the cardiac cycle. It follows the QRS complex and is usually of the same deflection. The absolute ventricular refractory period is represented by the peak of the T wave. At this point, the cardiac cells are repolarizing, or restoring, their electrical charges. No transmission of an electrical impulse can occur.
A U wave is seen occasionally, following the T wave. This wave is not clearly understood. It is thought to represent electrolyte imbalance.
The ST segment represents the time between depolarization of the ventricles and the beginning of ventricular repolarization. It is measured from the end of the S wave to the beginning of the T wave. Normally, this wave is isoelectric.
The interval between the P wave and the QRS complex is referred to as the PR interval. It is measured from the beginning of the P wave to the beginning of the Q wave or to the beginning of the R wave if a Q wave is absent. It represents atrial depolarization and AV conduction time, the time required for an impulse to travel through the atria and arrive at the Purkinje fibers. The normal range is from .12 seconds to .20 seconds.
Measured by the formula
![]() , this corrects for alterations in heart rate. The refractory period of the ventricles (the period in which one depolarization occurs and before another can occur) is reflected by the QT interval, the time between ventricular muscle depolarization—the QRS complexand ventricular muscle repolarization—the T wave. It is measured from the beginning of the QRS to the end of the T wave. The QT interval is normally less than half of the R to R interval.
, this corrects for alterations in heart rate. The refractory period of the ventricles (the period in which one depolarization occurs and before another can occur) is reflected by the QT interval, the time between ventricular muscle depolarization—the QRS complexand ventricular muscle repolarization—the T wave. It is measured from the beginning of the QRS to the end of the T wave. The QT interval is normally less than half of the R to R interval.
The mechanical events of cardiac contraction are influenced by four factors: