|
|
| Contents | Previous | Next |
One of the critical concerns in periodontology and dental implantology is the therapeutic management of new bone formation. The objective of this extensive study for many years has been the acquisition of a complete understanding of the nature of osteoblasts (bone-forming cells) and their lineage, the factors that exert regulatory control over osteoblast behavior and bone formation locally, and the manner in which these different regulatory mechanisms interact. Hopefully such an understanding will culminate in the development of novel biologically based therapies for the management of bone regeneration, in dental applications as well as in many other disciplines, such as orthopedic surgery, and maxillofacial and craniofacial surgery.17
Hughes et al.17 reviewed the literature on the nature of the osteoblast phenotype, its lineage and the genetic regulation of osteoblast differentiation and function; the role played by a wide variety of cytokines and growth factors in regulating osteoblast differentiation and function; the mechanisms by which some of these cytokine pathways may be regulated and interact with each other in the co-coordinated control of osteoblast function; and the pharmacological application of cytokines to achieve therapeutic management of bone formation locally in vivo.17
Hughes et al.17 described cytokines as “soluble, secreted glycoproteins which act as local signaling molecules to control and co-ordinate cellular behavior and function”. Cytokines function as ligands binding to cell-surface receptors, which then trigger serial intracellular signaling events, eventually leading to the modulation of gene expression in their target cells.17
Generally considered to be a subset of cytokines, growth and differentiation factors have anabolic physiological and pathophysiological roles in the regulation of tissue growth and healing. The terms ‘growth factors’ and ‘cytokines’ are used widely. Considering the different complex roles that these molecules may all have on cell and tissue function, the distinction between them is most likely an artificial one. In view of the protocol that guides the use of these terms, Hughes et al.17 have used the term ‘growth factors’ here to describe this distinct subgroup of cytokines. They have discussed the role of cytokines separately that are associated with inflammation in bone cell regulation.17
Osteoblasts are specialized cells derived from the mesenchyme; their function is the deposition and maintenance of skeletal tissues. Like any cell type, they are defined by the genes they express. These genes are a subset of the total genome and they define the unique function of the cells producing them. In particular, osteoblasts express the genes that code for bone matrix proteins as well as high levels of the tissue nonspecific form of alkaline phosphatase. Osteoblasts are also frequently characterized by their expression of parathyroid hormone/parathyroid hormone receptors, which are responsible for the classic cAMP response via stimulation with osteotropic parathyroid hormone.17
The bone matrix proteins consist of type I collagen and a number of other matrix proteins. To a greater or lesser degree these proteins are characteristic of bone or expressed specifically in it. Collagen type I comprises about 90% of the organic matrix of bone; it is responsible for the strength, structure and elasticity of mature bone tissue.17
Osteocalcin (bone GLA protein), osteopontin, osteonectin and bone sialoprotein are referred to as the noncollagenous proteins of bone. Moreover, the proteoglycans decorin and biglycan are major components of the bone organic matrix. Together, these proteins exhibit calcium-binding activity, which is thought to be responsible for the superbly intricate regulation of hydroxyapatite crystal deposition, orientation and specific crystal size present in the mineralized bone matrix. The Arg–Gly–Asp (RGD) peptide cell-attachment sequences are known to be present in bone sialoprotein and osteopontin which regulate attachment of both osteoblasts and osteoclasts to the bone matrix.17
Osteocalcin is considered to be the most bone-specific of the noncollagenous proteins of bone. In the mature organism, expression of osteocalcin is restricted to bone, dentin and cementum. In particular it is the data from mouse 'knockout' models that provides evidence that bone matrix proteins have a significant role in regulating the comprehensive control of bone mass. In particular, as its main phenotype, the osteocalcin knockout mouse exhibits an overall increase in bone mass.17
The expression of bone-specific genes and proteins provides valuable markers that give evidence of the bone osteoblast phenotype in vitro. Comprehending the transcriptional regulation of these genes is fundamentally important to understanding the regulation of osteoblast differentiation and function.17
Osteoblasts arise from multipotent mesenchymal stem cells. These stem cells produce progenitor cells of restricted osteoblast lineage that can go through a progression of proliferation or amplification stages before they express identifiable and specific osteoblastic markers. Early studies were conducted in 1967 and 1970 to determine the kinetics of normal osteoblast turnover using pulse-chase 3H-thymidine labeling. The results showed that osteoprogenitor cells were recruited from surrounding connective tissues toward the periosteal layer on the bone surface.17
These early studies described the periosteum as consisting of two layers: a fibroperiosteum and an inner osteogenic periosteum. The fibroperiosteum is located on the outside and most distant from the bone surface. The inner osteogenic periosteum is located where the pre-osteoblast is found; the pre-osteoblast is the first directly recognizable cell of the differentiating osteoblast lineage.17
While pre-osteoblasts retain a proliferative capacity, they can, at the same time, express many of the proteins associated with the mature osteoblast phenotype, including osteopontin and alkaline phosphatase. Lying immediately adjacent to the bone is the mature osteoblast layer containing a post-mitotic cell that expresses all of the characteristics of the mature phenotype. It is from these descriptions that researchers developed the model of osteoprogenitor cells proliferating from adjacent connective tissues which then go through further differentiation, ending with the expression of the mature phenotype. 17
The findings from these studies indicate that mature osteoblasts have an average lifespan of about 1 month. They then undergo apoptosis with 2 further options: they may be replaced by newly differentiated osteoblasts, or about one-third of these cells may be incorporated into deposited bone matrix as osteocytes.17
Therefore, the most differentiated cell of the osteoblast lineage is the osteocyte. It may persevere in bone matrix for long time periods. While some functions of the osteocyte remain a mystery, there is strong evidence that the osteocyte not only plays a vital role in maintaining bone mass, but it the primary bone cell responsible for mechanotransduction. Regular mechanical stimulation is necessary for the maintenance of bone mass.17 “Mechanotransduction refers to the many mechanisms by which cells convert mechanical stimulus into chemical activity.”19
Scientists believe that bones flex subtly in response to any loading which results in a pulsatile fluid flow through osteocyte canaliculi, with subsequent signal transduction by osteocytes for the purpose of retaining bone mass. The exact molecular mechanisms responsible for this are not yet fully understood. However, studies have demonstrated that mechanical strain and pulsatile fluid flow are able to induce osteocytes to produce signaling molecules such as nitric oxide in vitro. 17
Scientists have recently described a bone morphogenetic protein (BMP) inhibitor named sclerostin. Having been localized specifically to osteocytes, it may play a role in regulating overall bone mass. Similar pulse-chase 3H-thymidine-labeling experiments revealed that around the periodontal tissues there is a continual recruitment of proliferating cells from bone marrow stroma through vascular channels that communicate with the periodontal ligament.17
These cells are thought to exhibit preferential migration to osteoblast (and cementoblast) surfaces, eventually undergoing osteoblastic differentiation similar to the kinetics described in other bony tissues. This is consistent with the mechanism that recruits osteoprogenitor cells from stem cells located in the bone marrow stroma to the periodontal ligament. This process allows for constant repopulation of the periodontal ligament by relatively undifferentiated mesenchymal cells; and as such, the periodontal ligament is a rich source of progenitor cell that promote new bone formation.17
Thus the osteoblast lineage is comprised of mesenchymal stem cells that exhibit self-renewal. They may include cells that are multipotent and cells with more restricted potentials. It has been proposed that the transcription factor, TAZ, is the likely mediator in the process whereby these cells are committed to the osteoblast lineage, leading to committed monopotent osteoprogenitor cells with extensive proliferative capacity. Transcription factors, including Cbfa-1/Runx-2 and osterix, ultimately regulate the differentiation of mature osteoblasts.17
In the past decade enormous advances have been achieved in unlocking the secrets of cellular and molecular control of bone formation. The in vitro models of osteoblast differentiation and formation that were developed proved essential in understanding the effects of specific growth factors and growth factor-induced transcription factors on osteogenesis. Included in these models are the use of both primary cell cultures and established cell lines from a range of different species and with distinct characteristics.17
Interpreting the large amount of data from the in vitro models may sometimes be problematical. There can be variations in different model systems. The cells may manifest different responses depending upon the precise stage of target cells through the lineage at different stages of differentiation.17
In vitro, when osteoprogenitor cells are treated with fibroblast growth factor-2, the result is a mitotic response but the expression of osteoblast differentiation markers is suppressed. Paradoxically, when this same treatment is applied in vivo, it may result in a general stimulation of bone formation. Hughes et al.17 proposed the theory that fibroblast growth factor-2 stimulates the expansion of relatively undifferentiated cells. This is interpreted as a suppression of differentiation in vitro but eventually causes an increase in the number of mature osteoblasts in vivo.17
The goal of tissue engineering is the production of tissues that are identical, both structurally and functionally, to the original tissues they are to replace. Knowledge of the physiological role of different signaling molecules required for the process of bone formation is central to the emerging strategies for growing bone therapeutically. It involves the coordinated expression of many molecules to direct the formation of bone, such as growth factors, BMPs and specific transcription factors. These molecules appropriate developmentally derived signals for the purpose of inducing cellular and molecular stimuli to direct cellular commitment and differentiation in the correct spatial and temporal sequence.17
Researchers predict that the periodontal tissue engineering of the future may utilize suitable gene expression systems for the regulated delivery of regenerative growth factors within the proper biological scaffold. Growth factors are soluble proteins; they perform the function of signaling agents for cells. They utilize receptor–ligand binding to influence critical functions, such as cell division, matrix synthesis and tissue differentiation. Through experimental studies it is now accepted that growth factors play a variety of important roles in bone formation and bone repair.17
Growth factors found in bone, cementum and healing tissues (within the periodontal environment) include transforming growth factor-β, basic fibroblast growth factor, insulin-like growth factors, platelet-derived growth factor and BMPs. With the discovery of recombinant growth factor proteins, researchers have become interested in their potential therapeutic use to treat bone disorders (including periodontal disease). It is critical that we comprehend the nature of the biological characteristics of growth factors and their potential for periodontal tissue engineering as advances in technology enable them to be used as therapeutic agents.17
Scientists have outlined various candidate growth factors and their respective signaling pathways during bone formation. Yet, they have achieved only a minute understanding of the complex processes by which these growth factors interact to regulate bone formation and repair. Biologically it seems reasonable that multiple bioactive factors are involved in osteoblast development and bone formation, and that these factors most likely act in a sequential manner.17
They have also identified various growth factors and their downstream molecular targets during osteoblast differentiation. According to these findings, at least some of these growth factors or their genetic effectors may have the potential for therapeutic application in bone regeneration. Transforming growth factor-β, fibroblast growth factors, platelet-derived growth factor and insulin-like growth factor are known to affect osteogenic cells.17
Transforming growth factor-β is only one of the proteins belonging to this large superfamily of related proteins; also included are BMPs, growth and differentiation factors, activins, inhibitins and anti-Mullerian hormone. All members of this family participate in the important functions of regulating cell proliferation and differentiation, and producing extracellular matrix.17
Transforming growth factor-β is comprised of five isoforms: transforming growth factors-β1, β2, β3, β4, and β5. High levels of transforming growth factor-β are found in bone, platelets and cartilage, although most cells synthesize and respond to it. At the protein level, transforming growth factor-β1 is the most abundant isoform. Activation of transforming growth factor-β is a highly regulated process, but once activated, it readily interacts with transmembrane serine/threonine kinase receptors.17
According to Hughes et al.17 “Pivotal genetic studies identified that intracellular transforming growth factor-β signaling is mediated through mothers against dpp (Mad) in Drosophila melanogaster and subsequently the homologous genes in vertebrates (Smads) have been identified…” BMPs have also been shown to signal through the Ras/mitogen-activated protein kinase (MAPK)/activator-protein-1 (AP-1) pathway, although Smads are the primary mediators in the transforming growth factor-β signaling pathway.17
Transforming growth factor-β acts during the stages of early bone formation to both recruit and stimulate osteoprogenitor cells to proliferate and provide a collection of early osteoblasts. However, transforming growth factor-β blocks differentiation and mineralization in the later phases of osteoblast differentiation. Highly dependent on bone cell source, the dose applied, and the local environment, these effects may possibly be the consequence of inhibition of DNA synthesis at high concentrations of transforming growth factor-β. 17
In addition, transforming growth factor-β is known to inhibit the expression of the Runx2 and osteocalcin genes. Cbfa1/Runx2 controls the expression of these genes in osteoblast-like cell lines, and this is mediated by Smad3. The interaction of transforming growth factor-β with a variety of other growth factors in bone generates a complex response.17
Additional research is recommended to illuminate the role of transforming growth factor-β during periodontal regeneration and wound healing, with the goal of determining the inter-relationship between transforming growth factor-β and other growth factors that produce effects during different stages of osteoblast differentiation.17
Bone morphogenetic proteins are secreted as signaling molecules. They demonstrate various functions during development and cell differentiation. They recapitulate the entire sequence of events occurring during endochondral ossification, thus demonstrating their extraordinary ability to induce cartilage and bone formation from nonskeletal mesenchymal cells.17
Since their discovery, scientists have cloned and sequenced them, and manufactured recombinant proteins. Many in vivo and in vitro studies have focused on these recombinant proteins. Researchers are predicting that they will soon be employed therapeutically in orthopedics, craniofacial surgery and dentistry.17
BMPs, like transforming growth factor-β, are expressed both during embryonic development and well into adulthood. Researchers have identified more than 20 BMP-related proteins and reported diverse biological activity as the bone morphogenetic proteins bind to their specific receptors with different affinities. Bone morphogenetic proteins are known to bind to two distinct type I and II serine/threonine kinase receptors. Although both are necessary for signal transduction, it is the type I receptor that determines specificity of intracellular signals. Once the signals are activated, Smad-dependent and Smad-independent pathways transmit them intracellularly.17
Amidst the many BMP family members, it is BMP-2, BMP-4, and BMP-7 that demonstrate crucial roles during osteoblast commitment and differentiation. BMPs exert their primary effect on pluripotent cells that are able to differentiate into other mesenchymal cell types. BMP-2, BMP-4, and BMP-6 are all competent to direct these mesenchymal cells to commit to an osteoblastic pathway.17
With the formation of bone nodules and expression of markers of the mature osteoblast phenotype, bone morphogenetic proteins can increase the differentiation of committed cells to the osteoblast lineage. Gene-expression studies have revealed that a robust osteogenic response can be elicited by directly delivering an adenovirus with BMP-7 to gingival or dermal fibroblasts.17
From the perspective of both the basic molecular and potential therapeutic pathways, there has been a sudden increase of interest in potential molecular targets of BMPs in recent years. The result has been substantial insight into the genetic cascade that leads to bone formation. The transcription factors Cbfa1/Runx2, osterix, and TAZ are among these specific targets.17
Under certain conditions during osteoblast differentiation, bone morphogenetic proteins can upregulate Cbfa1/Runx2; thus, it is a potential downstream target of BMPs. However, Smad complexes are also capable of directly interacting with and activating target genes independently of Cbfa1/Runx2.17
There is emerging evidence to suggest that Wnt/β-catenin signaling may mediate some aspects of BMP-2-induced differentiation; although late osteoblast gene expression is activated by co-operation between β-catenin and Smads. Since β-catenin is the classical effector of Wnt protein signaling, scientists have suggested a role for this group of proteins in BMP-2 signal transduction.17
Fibroblast growth factors comprise a family of structurally related polypeptides. They are known to play a vital role in angiogenesis, (process of vascularization of a tissue involving the development of new capillary blood vessels3) and mesenchymal cell mitogenesis (the process of stimulating transit through the cell cycle3).17 Fibroblast growth factor proteins signal via membrane-spanning tyrosine kinases (FGFR) in order to mediate their range of effects. A wide variety of mechanisms exist for receptor regulation and availability.17
Abnormalities in ossification are associated with mutations in these receptors. Several craniosynostosis syndromes are caused by activating mutations in FGFR2 which affect the proliferation and differentiation of osteoblasts. Such events highlight a key role for these molecules in control of bone formation.17
Fibroblast growth factor-1 and fibroblast growth factor-2 are the most abundant proteins in normal adult tissues. Generally more potent than fibroblast growth factor-1, fibroblast growth factor-2 is expressed by osteoblasts, but other fibroblast growth factors are not nearly as universally expressed. Although fibroblast growth factor signaling is thought to be associated with bone development, research conducted on null mutant mice has not yet fully disclosed the function of this family in skeletal development.17
Fibroblast growth factor-1 and fibroblast growth factor-2 do not increase collagen production or alkaline phosphatase in differentiated osteoblasts, but they do stimulate osteoblast proliferation in vitro. However, these effects may be differentiation stage-specific considering that constitutive fibroblast growth factor signaling inhibits osteoblastic differentiation and noticeably increases apoptosis when cells are exposed to differentiating conditions.17
Not only are fibroblast growth factors strongly mitogenic to bone marrow stromal cells, they are also capable of maintaining self-renewal of these cells in culture. Fibroblast growth factor has an overall stimulatory effect on osteoblasts when the treatment is short term and followed by its removal from culture. Hughes et al.17 suggested that these growth factors, used as an adjunctive agent, may have the potential to increase formation of bone. They act primarily to stimulate proliferation in immature cells and concurrently inhibit differentiation, leading to expansion of the osteoblast progenitor pool. These findings are consistent with their general effects in vitro.17
Platelets secrete platelet-derived growth factor during the early phases of fracture healing, but it is present in a variety of tissues including bone. It is believed to act both systemically and locally since it is expressed by various tissues.17
Platelet-derived growth factor is comprised of two polypeptide chains; it contains two gene products (A and B), and exists in three different isoforms (AA, BB, AB) of these two gene products; these in turn bind to two separate a and b receptors. Platelet-derived growth factor (a powerful mitogen for connective tissue cells), is synthesized by and stimulates mesenchymal cells; nonetheless, it lacks the powerful properties necessary for bone-induction.17
The isoforms of platelet-derived growth factor exert a strong chemotactic effect on osteoblasts and other connective tissue cells. In addition they may possibly mobilize mesenchymal cells during bone development and remodeling. By upregulating collagenase transcription and increasing interleukin-6 (IL-6) expression in osteoblasts, platelet-derived growth factor may also directly and indirectly influence bone resorption.17
Other growth factors, such as transforming growth factor-β, exert paracrine regulation in osteoblasts, in addition to the osteoblastic autoregulation by platelet-derived growth factor. 17
Paracrine regulation is a “form of signaling in which the target cell is close to the signal releasing cell. Neurotransmitters and neurohormones are usually considered to fall into this category”.3
Generally speaking, in bone regulation, the primary physiological effects of platelet-derived growth factor are that of nonspecific mitogens. However, the effects of PDGF on cell chemotaxis and neovasculogenesis may be especially significant during would healing.17
The roles played by growth hormone and insulin-like growth factors are critical in skeletal development. Growth hormone helps regulate skeletal growth and stimulates target cells to release insulin-like growth factor. “Insulin-like growth factors are bound to binding proteins” and this serves as another crucial mechanism to control insulin-like growth factor activity.17
Scientists have identified two insulin-like growth factors: insulin-like growth factor-1 and insulin-like growth factor-2. Both exist in high concentration in serum. Although there may be inter- and intra-species differences, insulin-like growth factor-2 is more abundant in bone, but insulin-like growth factor-1 may be the more potent of the two. Researchers emphasize that the regulation of insulin-like growth factor is a complicated process, and in skeletal cells, the mode of action of growth hormone remains unidentified for the most part.17
All of the major hormones that regulate the skeleton have noteworthy effects on skeletal insulin-like growth factor, and so do many other growth factors, such as BMP-2, transforming growth factor-beta and fibroblast growth factor. Insulin-like growth factors increase osteoblast proliferation and have a significant role in stimulating the function of mature osteoblasts.17
Similar to other growth factors discussed here, the response of osteoblasts to insulin-like growth factor signals may be influenced by both the differentiation status of the cell and cell type. Insulin-like growth factor-1 upregulates osterix, which is the osteoblast-associated transcription factor at the molecular level, but it does not upregulate Cbfa1/Runx2. In combination with BMP-2, insulin-like growth factor-1 acts synergistically on osterix expression.17
The scientific community broadly accepts the decisive function that insulin-like growth factors have in bone remodeling, yet they are uncertain of the actual role that they play. They do agree that it is important to understand this role in the context of the complex inter-relationships formed by the components of the insulin-like growth factor system occurring in vivo. In general, the research findings indicate that the foremost effects of insulin-like growth factors are to promote osteoblastic late-stage differentiation and activity.17
Wnt proteins comprise a family of highly conserved signaling molecules that are secreted, and during embryogenesis, they regulate cell-to-cell interactions.18 They also help to regulate normal physiological processes in adult animals.19 Both Wnt genes and Wnt signaling are linked to cancer.18
“The name Wnt was coined as a combination of Wg (wingless) and Int.” Originally identified as a segment polarity gene in Drosophila melanogaster, the wingless Wg gene functions during embryogenesis and also during adult limb formation during metamorphosis.19 “The INT genes were originally identified as vertebrate genes near several integration sites of mouse mammary tumor virus.”19
With a common evolutionary origin evidenced by similar amino acid sequences of their encoded proteins, the Int-1 gene and the wingless gene were found to be homologous.19 “As a major class of secreted morphogenic ligands of profound importance, Wnts help to establish the pattern of development in the bodies of all multicellular organisms studied. Some components of the signaling pathway are remarkably conserved in a wide variety of organisms, from Caenorhabditis elegans to humans. Protein homology suggests that several distinct Wnt ligands were present in the common ancestor of all bilaterian (bilaterally symmetric) life, and certain aspects of Wnt signaling are present in sponges and even in slime molds.”19
Evidence is increasing to support the role of Wnt signaling in the regulation of early stages of the osteoblast lineage. Comprised of over 15 related extracellular signaling molecules, Wnts show ligand binding with their receptor, Frizzled, and co-receptors LRP5/6. Ligand binding stabilizes the protein β-catenin by stimulating a series of intracellular reactions. β-catenin is subsequently translated to the nucleus, there it acts as a co-factor with other DNA-binding proteins to regulate gene transcription.17
It is believed that Wnt proteins bind to receptors of the Frizzled and LRP families on the cell surface.18 The signal is transduced to β-catenin via several cytoplasmic relay components. β-catenin subsequently enters the nucleus and forms a complex with TCF to activate transcription of Wnt target genes.18
Although there is evidence for Wnt signaling acting through the induction of BMPs, and, conversely, BMPs acting through the induction of Wnt signaling in mesenchymal cells, the relationship between Wnt signaling and other growth factor-mediated osteoblast effects is currently perplexing. Even though presently there is minimal to no information available about the role of Wnt signaling in local bone formation during wound healing, scientists believe that this signaling pathway probably plays an important role in the early events of local bone formation.17
Numerous other growth factors are capable of interacting with cells of the osteoblast lineage and possibly function in the local regulation of bone formation. Although epidermal growth factor receptors may be downregulated in mature osteoblastic cells, osteoprogenitor cells do respond to epidermal growth factor stimulation. When in vitro cell cultures are treated with epidermal growth factor, cell proliferation increases and bone module formation is suppressed.17
Epidermal growth factor utilizes numerous signal transduction pathways commonly used by platelet-derived growth factor. However, subtle differences in intracellular signaling have recently been reported in these two signaling mechanisms when using a proteomic approach.17 In proteomics one analyses the expression, localizations, functions, and interactions of the proteins that are expressed by the genetic material of an organism.3
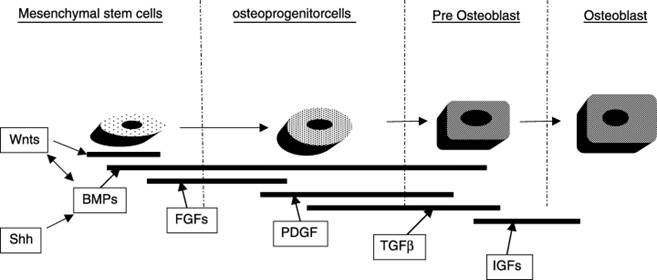
Another growth factor-like protein, sonic hedgehog (Shh), binds to a specific cell-surface receptor, patched (Ptc). Shh is known to induce osteoblastic commitment in cultured multipotent mesenchymal stem cells and to promote differentiation in osteoblasts. BMP antagonists can block these actions, indicating that “Shh is an upstream regulator of BMP production and the Shh-activated intracellular signaling molecule, Gli, can upregulate BMP expression by direct interaction with bmp gene promoter”.17 See Figure 2.
|
|
Enamel matrix derivatives (EMD), the major component of commercially available Emdogain®, are presented for comparison of their biological activity with growth factors discussed above. The main biological effects of EMDs are ascribed to their predominant protein, amelogenin; however, the remaining fraction comprises less well characterized factors. Not a classic growth factor, amelogenin is a cell-adhesion matrix-bound protein.17
Scientists have proposed that specific amelogenin gene products probably have activity as epithelial-mesenchymal signaling molecules. Research has also indicated that the leucine-rich amelogenin peptide, an alternate splice variant of the amelogenin gene, may also act directly to signal cementoblasts, and by implication, osteoblasts also.17
Clinicians have used enamel matrix derivative to regenerate periodontal tissues. The exact regenerative mechanism remains elusive and investigators have ascribed its therapeutic effectiveness to various substances: amelogenin, non-amelogenin enamel matrix proteins, and growth factors. In vitro studies have demonstrated that EMDs enhance matrix production, proliferation and bone nodule formation in cultures of periodontal ligament cell, as well as increase differentiation of human and murine osteoblast cell lines. In some osteoblast cell lines, they stimulate phenotypic bone markers.17
Some studies indicate that EMDs have no effect on osteoblastic differentiation, although they stimulated other growth factors. Furthermore, both transforming growth factor-β and BMP-like growth factors have been found in noncommercial EMDs. In the search for downstream target genes, scientists have discovered EMD response elements in the gene promoter for rat bone sialoprotein that may help to regulate EMD effects on transcription of the gene for bone sialoprotein.17
Utilizing a cDNA microarray study, investigators examined EMD-mediated changes in gene expression in periodontal ligament cells in vitro. They discovered the downregulation of genes involved in the early inflammatory phases of wound healing, with the concurrent upregulation of genes encoding molecules that promote growth and repair.17 “cDNA is the acronym for “complementary DNA that is synthesized from a messenger RNA template, the single-stranded form is often used as a probe in physical mapping to locate the gene or can be cloned in the double stranded form. Viral reverse transcriptase can be used to synthesize DNA that is complementary to RNA (for example an isolated mRNA)”.3
When cementoblasts were treated in vitro with EMDs, osteocalcin expression decreased, while osteopontin expression increased. In general, data confirms the positive effect of EMDs on osteoblast differentiation; however, scientists need further studies to determine which molecules in EMDs stimulate osteogenesis and to elucidate their exact modes of action.17
In summary, substantial progress was made when scientists were able to describe how growth factors regulate osteoblasts and some of the molecular mechanisms that contribute to these processes. At times the data can be both complex and contradictory; yet, there emerges an overall pattern of a rigorously regulated, temporal sequence of coordinated growth factor expression that can be regulated at any stage of the differentiation cycle.17
Not only do osteoblasts respond to growth and differentiation factors, they also respond to various cytokines that act locally. These cytokines are molecules that control the behavior of bone cells during inflammation, infection and wound healing. The response of these cells may be both anabolic and catabolic. This section concentrates on the capacity of the major pro-inflammatory cytokines to regulate osteoblast differentiation and function and finally introduces the link between the osteoblast and regulation of bone resorption.17
Interleukin-1 is described as a multipotent cytokine because it has the ability to produce or influence several effects or results. It is comprised of two individual peptides, IL-1a and IL-1β, which demonstrate similar biological activities. IL-1 is produced by both hematopoietic and mesenchymal/osteoblastic cells. Research findings have confirmed that IL-1 regulates both resorption and formation of bone. However, the results from different studies diverge when it comes to the effects of IL-1 on osteoblast function.17
For example, the presence of enhanced alkaline phosphatase activity and bone nodule formation demonstrate that both IL-1a and IL-1β inhibit osteoblast proliferation and enhance bone formation. Conversely, IL-1a and IL-1β may stimulate osteoblast proliferation, including DNA and protein synthesis, while inhibiting bone formation and osteocalcin and type I collagen production, contingent upon the differentiation stage of the cell, prolongation of the culture period, and concentration of the cytokines.17
Hughes et al.17 reported that interleukin-1 can also stimulate osteoblasts to produce other pro-inflammatory cytokines, such as IL-6, IL-7 and tumor necrosis factor-a (TNF-a), as well as other inflammatory mediators, such as prostaglandin E2 and nitric oxide.17
Recently it has become apparent that osteoblasts enjoy a comprehensive role in orchestrating the remodeling process of bone. Not only is the function of osteoblasts not restricted exclusively to bone formation, it is now accepted by the scientific community that osteoblasts actually initiate bone resorption. Apart from forming the mineral and organic extracellular compartment of bone, the osteoblast, in cellular terms, provides essential and sufficient stimuli via cell–cell interaction to control the behavior of the osteoclast.17
Two factors produced by osteoblasts/stromal cells, M-CSF and RANKL, are the molecular determinants of this interaction. M-CSF is secreted and RANKL is primarily cell-membrane bound. The binding of M-CSF to c-Fms on the surface of osteoclast precursors enhances their proliferation and survival. RANKL is a member of the TNF ligand superfamily. It is an absolute requirement for osteoclast formation. RANKL is definitely the factor that stimulates the osteoclast precursors to differentiate into mature osteoclasts and activates bone resorption.17
Osteoprotegerin (OPG) is a soluble nonsignaling decoy receptor and a member of the TNFR superfamily with high homology to RANK. OPG can block the effects of RANKL. Osteoblasts and stromal cells secrete OPG, which when bound to RANKL prevents RANKL/RANK interaction and, as a result, the downstream events.17
Mice deficient in RANKL lack functional osteoclasts and develop osteopetrosis (formation of abnormally dense bone3). On the other hand, mice deficient in OPG develop early-onset osteoporosis (pathological decrease in the amount of bone mass, leading to fractures after minimal trauma3). This illustrates the indispensable role played by a balanced RANKL/OPG expression in the remodeling of physiological bone.17
Studies have revealed an increase in the expression ratio of RANKL/OPG in diseases characterized by bone loss associated with inflammation, including rheumatoid arthritis and periodontitis. IL-1, IL-6, and TNF-a all potently induce osteoclast differentiation and bone resorption. Scientists have proposed that the catabolic effects they have on bone are largely mediated not only through osteoblasts/stromal cells, but even more explicitly by regulation of the expression of RANKL and OPG in these cells.17
Scientists have reported that a number of other cytokines also regulate bone cell behavior. Interleukin-7 and IL-10 inhibit osteoblast differentiation. Not only do interleukin-4 and IL-13 inhibit osteoblast differentiation, but mice that over-express IL-4 demonstrate decreased bone formation due to a bone-lining osteoblast reduction in number and activity. 17
Structurally and functionally homologous to Interleukin-1, IL-18 utilizes differential signal-transducing pathways and preferentially activates p38 MAPK instead of NF-?B. Produced by osteoblasts/stromal cells, it acts as a mitogen to them and inhibits the differentiation of osteoclasts.17
Produced by activated T cells, interferon-? is a lymphokine that powerfully inhibits osteoclastogenesis, additionally inhibiting osteoblast proliferation and synthesis of collagen and noncollagen proteins.17
To summarize, inflammatory cytokines probably exert both anabolic and catabolic effects on osteoblast function, and as a result, on bone physiology.17 Unfortunately there is minimal information on how these anabolic and catabolic effects could interact directly with growth factor pathways. Hughes et al.17 emphasize that these interactions are likely to be extremely significant.
While researchers have demonstrated that both IL-1 and TNF-a may modulate the effects of BMPs on osteoblasts, they have not explored the possible mechanism of these interactions. Consequently, they must consider the putative cellular responses elicited by cytokines upon osteoblasts during inflammation when undertaking any tissue engineering approach that utilizes these cells for bone regeneration.17
This is a legitimate concern in general and not only in the instance of a direct treatment strategy which administers exogenous cytokines. It is quite likely that various other primary molecular agents may stimulate target cells to produce secondary inflammatory responses, which consequently impact bone cellular dynamics in an autocrine or paracrine fashion.17
This subject is especially important in the case of catabolic effects, which can either inhibit bone formation, or, most commonly, enhance bone resorption. Effectively addressing the potential side-effects of these cytokines may help to promote the bone-remodeling balance that would favor bone formation, with the ultimate goal of achieving clinical bone regeneration.17
Most of the current knowledge about the mechanisms of action of growth factors and cytokines on their target cells has come from in vitro studies, or occasionally from transgenic and null mutant (knockout) mouse models. The advantage of these methodologies is the powerful manner in which they dissect out specific signaling pathways because of the opportunity to control many of the extrinsic variables. This enables investigators to study the role of individual specific molecules and signaling pathways.17
Whereas in vivo cells are simultaneously exposed to a broad collection of these signaling pathways, it is the total sum of these interactions that produces the vigilantly coordinated control of the osteoblastic function in vivo. The goal is to present a complete picture of the complex interactions in cytokine signaling pathways during bone growth. At the present time scientists can provide only examples of such interactions because only partial data is available on the interactions of different pathways during the coordinated regulation of bone growth.17
Various mechanisms have been identified that can regulate different growth factor pathways. The final observed outcome can be fine tuned by the interactions of different signaling pathways with each other. The mechanisms are: “1. The regulation of growth factor activity by the secretion of soluble antagonists which are often regulated in both autocrine and paracrine fashions, 2. The regulation of receptor expression on specific cells, and 3. The interaction between intracellular signaling mechanisms in both antagonistic and synergistic ways.”17
In theory, the pharmacological application of growth factors provides a local single dose of the factor. The concentration of growth factors in this single dose may be thousands of times higher than that normally seen physiologically.17 If the normal regulatory feedback mechanisms which control that factor are temporarily overwhelmed by such a large dose, Hughes et al.17 reported that the pharmacological effects of the growth factor may be quite distinct from its physiological activity.
Preclinical studies support this idea that the therapeutic application of any one of a number of growth factors could influence the normal bone-forming process in a positive way. For example, considering what is currently known about physiological function, “BMPs may increase osteoblastic stem cell recruitment and commitment, fibroblast growth factor may stimulate early progenitor cell number expansion, platelet-derived growth factor may also increase progenitor cell recruitment and proliferation, and insulin-like growth factors might act to increase final osteoblast differentiation and function”.17 Any of these actions may increase bone formation which is the desired final outcome.17
Achieving sufficient substantivity (duration of contact) for the factor to provide a continual therapeutic dose over a lengthy period of time is probably one of the main critical determinants of success for these treatments. Not much data has been published on the pharmacokinetics of growth factor delivery.17 However, Hughes et al.17 reported on a study in which platelet-derived growth factor with insulin-like growth factor-I was delivered in a collagen matrix. It had a half-life of <4 hours and lost activity completely by 96 hours.
To address the issue of half-life, investigators explored a number of different delivery systems for these applications, including collagen matrix, cross-linked gelatin pellets, calcium phosphate ceramics, and poly-lactic-acid (PLA)-based resorbable polymers. In an effort to provide continuing growth delivery factor, scientists tested somatic cell gene transfer of platelet-derived growth factor both in vitro and in animal models.17
While this approach might be able to achieve a sustained active concentration of growth factor to the site being treated, there are serious concerns about the safety of the adenovirus vectors used for transfer genes, in addition to other ethical and practical issues that must be surmounted before this could be accepted as a practical and clinically appropriate technique.17
Hughes et al.17 discussed various other studies that have tested the potential of platelet-derived growth factor and also with platelet-derived growth factor combined with insulin-like growth factor-1 or dexamethasone to stimulate tissue regeneration. When compared with controls, these animal studies generally revealed an enhanced rate with an increased total amount of regenerated tissue. However, there was only minimal evidence that insulin-like growth factor-1 was useful, either alone or in combination with platelet-derived growth factor.17
More recently, there was an interesting report of a large multicenter (180 patients) human study with platelet-derived growth factor. When compared with the carrier-only controls, group treated with recombinant human platelet-derived growth factor-BB with a β-tricalcium phosphate carrier showed improved bone fill and an increased rate of clinical attachment gain.17 Hughes et al.17 reported that while this is probably the first large human study to investigate the role of recombinant growth factor therapy, these promising results will need to be evaluated further over time.
Researchers have extensively tested bone morphogenetic proteins for their pharmacological suitability to increase new bone formation. In critical-size segmental defects in animals, bone morphogenetic proteins have demonstrated the potential to vigorously promote bone formation. They manifested evidence of their ability to regulate bone and periodontal regeneration.17
Animal models have been used to evaluate the ability of fibroblast growth factor-2 to promote periodontal regeneration. Fibroblast growth factor-2 demonstrated promising results in stimulating bone and periodontal regeneration in surgically created defects. Compared to the effects of other growth factors, the results from the initial studies of transforming growth factor-β were disappointing.17
The use of platelet-rich plasma has also generated clinical interest. Obtained from a patient's own platelets, it is an autologous preparation rich in a number of growth factors, including platelet-derived growth factor and trefoil factor family-β. Platelet-rich plasma has been applied clinically in the treatment of both periodontal disease and implants. Unfortunately, very limited suitable scientific evidence exists to evaluate the effectiveness of this procedure. Well-controlled clinical studies of platelet-rich plasma would be beneficial in determining whether this treatment has a potential comparable to the pharmacological use of recombinant growth factors.17
In conclusion, the sudden increase of knowledge and the new understanding of the role played by growth factors, their mechanisms of action and molecular signaling pathway discussed here, propose the potential for many new therapeutic targets, not only in the application of growth factors, but for the future use of growth factor inhibitors or agents that target specific areas in the intracellular signaling pathways.17
The monumental challenge remains to adapt knowledge from the basic science of bone cell physiology to therapeutically efficacious techniques for the future. It is the hope of the scientific community that these novel approaches will provide qualitative advancement in clinical outcomes compared to the techniques currently available.17