|
|
| Contents | Previous | Next |
Without growth factors, healing could not occur. According to Takata8 the complete restoration of the structure and function of tissues damaged by periodontal disease remains the objective of periodontal therapy, yet, it is very difficult to accomplish. Recent advances in periodontal wound healing concepts are encouraging. Complete restoration of the structure and function of damaged periodontal tissues becomes even more challenging when deep subcrestal periodontal tissues are involved.8
This ultimate goal may possibly be achieved by the following:8
Healing, which is divided into regeneration and repair, is a response to different types of injury en route to the re-establishment of normal structure and function. “Regeneration is the complete restoration of damaged tissues by proliferation and differentiation of the original parenchymal elements.”8 Repair, which is incomplete regeneration, achieves healing by replacing lost tissue with a nonspecialized connective tissue called granulation tissue that matures into a fibrous tissue and finally a scar.8
Even though nearly all connective tissue contains fibrillar elements, the term fibrous usually refers to tissue laid down at a wound site that is well vascularized initially at the granulation tissue stage, but later becomes avascular and is dominated by collagen rich extracellular matrix and scar formation.3 Clearly, regeneration is the more advantageous healing process. Although it is well documented that newts can regrow lost extremities, in general, we cannot expect regeneration in mammals, except where there is very minute tissue loss with the framework of the tissue intact.8
Cell proliferation is an absolute prerequisite for tissue restoration. Cells are categorized into groups based on their mitotic activity, i.e., whether they are labile cells, stable cells, or permanent cells. Even in the normal physiologic state, labile cells go through continuous replication.8
Examples of labile cells are epithelial cells found in the skin and mucosa, hematopoietic cells, lymphoid cells, etc. Stable cells experience decrease or loss of mitosis in the adult cell stage but are able to re-enter the cell cycle if the situation warrants. Stable cells include glandular parenchymal cells of the liver, pancreas, endocrine glands, etc. Permanent cells such as cardiac myocytes and neurons surrender their ability to proliferate. Tissue that is comprised of labile and stable cells can regenerate. However permanent cells cannot be renewed once they are lost.8
Traditionally, wound healing is divided into two types based upon on the type of wound. In healing by primary intention, the wound edges are apposed and there is minimal tissue loss. Healing by secondary intention is characterized by edges that are separated with a more extensive tissue loss. Healing by primary intention results in a small scar, whereas healing by secondary intention is followed by a large scar. The basic healing mechanism of wound contraction, repair, and regeneration is the same for both healing by primary and secondary intention. The differences are quantitative.8
Contraction is the mechanism that reduces the wound size with the help of myofibroblasts. Repair occurs through hemorrhage and clotting, organization via granulation tissue, and fibrosis with the appearance of a scar in the mature tissue. The manner in which collagen fibers are arranged in a wound is dependent for the most part on the direction of the tensile strength in the damaged tissue. When the collagen fibers in a scar are arranged functionally, it is defined as a functional scar.8
Even in mammals, it is possible for regeneration to occur to some degree. Injured superficial epithelium can be entirely restored, in the case of cutaneous and mucosal wound healing, by the migration and proliferation of cells from the wound margin. The epithelial cells that are regenerating migrate between the clot and the organizing granulation tissue until they come into contact with one another. They then experience contact inhibition which prevents further migration.8
In addition researchers have shown that the proliferation, migration, and differentiation of parenchymal and mesenchymal cells in healing, are regulated by a number of growth or differentiation factors and extracellular matrix components. Examples of growth or differentiation factors include epidermal growth factor, fibroblast growth factor, platelet-derived growth factor, macrophage-derived growth factor, vascular-endothelial growth factor, insulin-like growth factor, transforming growth factor, etc. Examples of extracellular matrix components are fibronectin, collagens, proteoglycans, laminin, etc.8
Periodontal tissues comprise a distinctive arrangement in which epithelial, soft and mineralized connective tissues collectively form a dentogingival junction. This dentogingival junction is an intricate structure, and maintenance of its integrity is essential in order to preserve the periodontal ligament underlying bone. Regrettably this junction is destroyed by the chronic inflammatory process related to periodontal diseases. In order to control the destructive effects of chronic periodontal diseases and to regenerate the lost tissues to some degree, the dentogingival junction would have to be restored.9
Whether surgical or nonsurgical techniques are selected in the conventional approach to periodontal therapy, the instrumentation used in debridement and surgery results in a wound in the already inflamed dentogingival tissues. The end result is that these therapeutic measures are mainly dependent upon the cellular and molecular processes necessary for wound healing. Numerous cellular and molecular events that are part of the healing process for periodontal wounds are analogous to those seen in wounds located in the rest of the body, with the exception that there is a mineralized tissue located at the junction of epithelium and connective tissue in periodontal wounds.9
After the periodontal surgical procedures, the tissues are considered surgically wounded sites; there is a subsequent surge of cellular and molecular events that initiate wound repair. In the past 20 years especially, many discoveries have contributed to our understanding of the biology of wound healing. Currently, progress in understanding the biology of periodontal tissues has provided us with surgical procedures that promote regeneration of periodontal tissues lost as a result of chronic inflammation.9
The principle of guided tissue regeneration as applied to the surgical treatment of periodontal lesions is an excellent example of the application of cell biology to improve the clinical outcome of therapy. While it is important to review current basic concepts of wound healing in general and contrast them to the concepts of periodontal wound healing, no distinction is necessary between the healing in different types of periodontal defects as the biological principles of wound repair would virtually be the same. This review is intended to enable the reader to extrapolate basic wound healing principles to the various periodontal surgical wounds.9
Wound healing is initiated by a temporary repair as a clot forms in the wounded tissues. Inflammatory cells invade the clot first and, followed by fibroblasts and endothelial cells, form granulation tissue. Next, the migration of epithelial cells covers the denuded tissue surfaces or forms a junction at the tooth-tissue interface. Lastly, maturation of the healing tissue matrix is accompanied by contraction or scarring.9
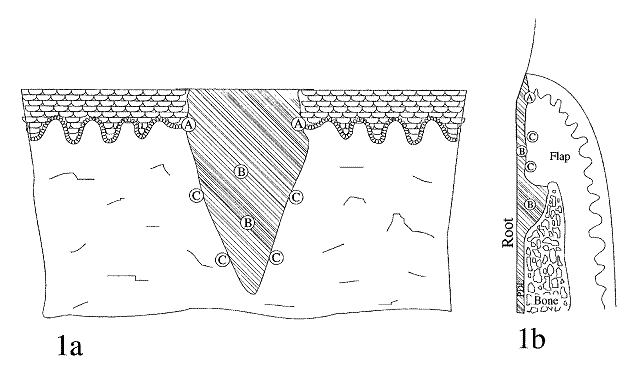
These wound healing events do not occur in isolated phases or strict sequence and overlap to some extent in time. This explanation is an oversimplified summary of the wound healing events, but it does reflect the important cellular and molecular mechanisms that have come to light in the past 20 years that may facilitate wound healing in the future.9 See Figure 1.
|
|
Cells from the labile or stable cell group comprise the periodontal tissues: gingiva, periodontal ligament, alveolar bone, and cementum. When periodontal tissues are injured, it is anticipated that these labile or stable cells will demonstrate their proliferative capability.8
The healing of periodontal tissue is accomplished by wound contraction, while repair is accomplished by the laying down of granulation tissue and regeneration. Regeneration occurs in the gingival epithelium and the localized areas of the wound margins; but since the framework of the periodontal tissues is usually destroyed by inflammation or surgical treatments, complete restoration of injured periodontal tissues is difficult to achieve. Since the tissue elements participating in the healing of the two areas are different, it is convenient to review the healing of the supracrestal gingival tissue involved in gingivitis first and then the healing of the supracrestal and subcrestal periodontal tissues involved in periodontitis.9
Gingivitis is described as the inflammation that is limited to the gingival tissue and it is demarcated from the deep subcrestal periodontal tissues by crestal and transseptal fibers. As a consequence, tissue damage is confined in the gingiva, and there is less damage to the fiber arrangement that hinders the down growth of epithelium over the cementoenamel junction. In order for the tissues damaged by gingivitis to heal, the injured gingival epithelium and connective tissues must be restored via periodontal therapy.8
If gingiva is removed by surgical excision, granulation tissue and epithelial cell migration from the adjacent mucosal tissue achieve nearly complete restoration. The granulation tissue rebuilds the majority of the gingiva in which fibers are arranged functionally in the direction of their tensile strength resulting in a functional scar. Epithelial cells migrate across the granulation tissue. Epithelial migration is normally inhibited by cell to cell contact, but in the periodontal wound there is no encounter of migrating epithelial cells on the surface of a tooth.8
If the aggregation of fibers is injured, then the regenerating epithelial cells migrate down into the deep periodontal tissue. In gingivitis, fortunately, the group of fibers physically blocks the down-growth of epithelial cells, and epithelial growth terminates apically in the area of the cementoenamel junction. Depending on their environment, the regenerated epithelial cells differentiate, respectively, into the junctional, oral sulcular, and oral gingival epithelium. Hence, the gingival tissues associated with gingivitis are capable of almost complete regeneration.8
When periodontal tissues have been damaged by periodontitis, the healing is more complicated compared to that of the more simplified healing in gingivitis. There are four different types of cells that contribute to the healing of periodontal tissues that have been damaged by periodontitis or surgical treatments such as flap procedures. These cells come from the gingival epithelium, gingival connective tissue, bone, and periodontal ligament.8
In 1976, a theory was proposed regarding the potential for periodontal tissue to repair itself. It suggested that cells that are first to repopulate the exposed root surface are the ones to establish the nature of the attachment that develops. Thus, the potential for repair may depend on which cell type wins the race to populate the root surface first.8
If epithelial cells populate the root surface first, specifically growing on the surface of the gingival granulation tissue, a long junctional epithelium will be the consequence. If gingival connective tissue cells populate the root surface first, the attachment will be one of connective tissue adhesion, and root resorption is likely to occur. If bone cells are the first to proliferate and make contact with the tooth surface, root resorption and ankylosis are the result. If periodontal ligament cells are the first to migrate and cover the denuded root surface, then new connective tissue attachment forms and this is the ideal attachment.8
Of these four healing patterns, proliferation of new connective tissue by periodontal ligament cells and new bone formation constitute regeneration. The other healing patterns are associated with repair. Clinical studies have demonstrated that new connective tissue attachment can be achieved by new attachment procedures including bone grafting. Further clinical evaluation demonstrated that the healing which follows new attachment procedures resulted in formation of a long junctional epithelium and no connective tissue attachment.8
Their conclusion was based on a histometric evaluation of connective tissue attachment levels that developed after four different regenerative procedures: 1.Modified Widman flap (gingiva is reflected to provide access for scaling and root planing, scalloped internal beveled incision to excise inflamed periodontal pocket wall, heals by long junctional epithelium); 2. Flap procedure in combination with transplantation of frozen autogenous bone; 3. Flap procedure in combination with implanted beta tricalcium phosphate; 4. Periodic root planing and soft tissue curettage.8
Epithelial cells migrate very swiftly and certainly faster than connective tissue cells, and there is no physical barrier to hamper the apical migration of epithelial cells. It naturally follows that the epithelial cells from wound margins would proliferate on the surface of granulation tissue resulting in a long junctional epithelium. If the tissue framework is not severely damaged, regeneration may occur in very limited areas at wound margins. Yet, after conventional periodontal treatments, healing results in repair not regeneration.8
Later studies illustrated the repair potential of periodontal tissues in another series of experiments. They demonstrated that cells originating from periodontal ligament are the only cells with the ability to produce a new connective tissue attachment. In the 1980’s various histologic and clinical studies supported the theory that new connective tissue can attach to a previously exposed root surface if cells derived from the periodontal ligament are stimulated to proliferate.8
When blood vessels are injured during therapeutic procedure such as surgery or periodontal treatment, extravasation of blood results.9 Extravasation refers to the leakage of blood from a blood vessel into the tissues.3 Blood coagulation and platelet aggregation form a fibrin-rich clot that occludes the injured blood vessel and temporarily protects the denuded tissues. The clot is comprised of platelets located in a complex of cross-linked fibrin fibers together with plasma fibronectin, vitronectin and thrombospondin.9
The clot performs various vital functions. Significantly, it acts as a reservoir of growth factors and cytokines. Degranulation of activated platelets releases the growth factors and cytokines from the clot to serve as a provisional matrix for cell migration. The growth factors and cytokines located in the fibrin clot may provide the signals that initiate wound repair.9
First, inflammatory cells are recruited to the wound site and then succeeded by epithelialization, granulation tissue formation and angiogenesis. Initially, the signals found in the clot usually recruit neutrophils and monocytes. Neutrophils perform the vital function of cleansing the wound of bacteria, foreign particles, and debris.9
It is significant that neutrophils perform the very important function of eliminating bacterial debris by releasing enzymes and toxic oxygen products. Not only do these enzymes and oxygen products kill bacteria but they also damage host tissues. Therefore, the greater the number of contaminating bacteria in the wound, the greater the potential there is for neutrophil-mediated destruction of host tissues.9
Neutrophils also produce pro-inflammatory cytokines, and their signals activate adjacent fibroblasts and keratinocytes. After a few days, the infiltration of neutrophils terminates, and they are ultimately phagocytosed by either macrophages or fibroblasts. Continuing recruitment of peripheral blood monocytes into the wound site is promoted, and upon activation, they become macrophages. Both fibrin and fibronectin in the clot function as a provisional matrix, supporting the inflow of monocytes and fibroblasts.9
Macrophages continue to provide for the phagocytosis of bacterial, cellular and matrix debris in the wound. Macrophages also maintain synthesis of growth factors and cytokines and secrete them into the wound locale. Thus, it is the macrophages that maintain the wound repair signals that were initiated by degranulating platelets and neutrophils.9
However, there is a lag time between wounding and the initiation of epithelial migration. Normally, the basal layer of epithelium attaches to the basal lamina in healthy gingival tissues. Keratinocytes have receptors on their surface called integrins. They use these integrin receptors to bind to laminin in the basal lamina.9
Integrins comprise a family of cell adhesion receptors; their function is to mediate cell surface interactions with extracellular matrix and sometimes with other cells.9 Extracellular matrix is any material produced by cells and secreted into the surrounding medium, but the term usually refers to the noncellular portion of animal tissues.3 This family of cell surface receptors has about 20 members.9
Each integrin receptor is comprised of a heterodimer made up of one a subunit and one b subunit in a non-covalent complex. Alterations in the combinations of various a and b subunits provide the integrins with specificity for the ligand.9 A ligand is any molecule that binds to another, usually referring to a soluble molecule such as a hormone or neurotransmitter that binds to a receptor.3
The migration of cells is fundamental to wound healing; therefore, it is essential to take into account how cells migrating into the wound change the expression of integrins receptors. In normal, healthy tissues, keratinocytes utilize the a6b4 integrins to bind to laminin found in the basal lamina. These a6b4 integrins possess intracellular links with keratin in the cytoskeletal network.9 Before keratinocytes can begin to migrate, they must lay the groundwork to facilitate this process. Those located at the edge of surgical wounds begin by dissolving the hemidesmosomal attachment. They then initiate expression of other integrins more appropriate to the wound environment.9
These migrating keratinocytes begin to express integrins α5β1 for binding to fibronectin, αVβ6 for binding tenascin, integrins αVβ5 for binding to vitronectin, and lastly they rearrange the distribution of integrins α2β1, a collagen receptor. Thus, keratinocyte mobilization and expression of new integrins promotes adhesion of the keratinocytes to matrix molecules in the provisional matrix and in the adjacent wound debris. It may also account for the lag time between wounding and the initiation of epithelial migration.9
The general consensus is that the basal cells contribute the majority of migrating keratinocytes, although there is data to suggest that some of the suprabasal cells may migrate also. After the epithelial cells have begun to migrate, basal layer cells close to the wound edge undertake proliferation, yielding an additional source of basal cells. The basal cell layer is characterized by an apical-basal polarity, which is lost at the wound edge, and then replaced by pseudopodia which extend from their free basolateral sides into the wound.9
Little is known about the driving biological force behind epithelial cell migration, but chemotactic factors, active contact guidance, absence of neighboring cells or combinations of the above are all proposed mechanisms. A compelling note of interest is that epithelial cell migration does not seem to be dependent upon cell proliferation, as is evidenced by the fact that molecules such as transforming growth factor-b promote the migration of epithelial cells in organ cultures, even though it is a potent inhibitor of keratinocyte proliferation.9
After the process of re-epithelialization is complete, the basal lamina components are laid down sequentially beginning from the wound margin, and the epithelial cells change back into their normal phenotype. Various growth factors appear to be significant in modulating the proliferation of keratinocytes in healing wounds. Notably, epidermal growth factor, transforming growth factor-alpha, heparin-binding epidermal growth factor and keratinocyte growth factor are among the significant growth factors.9
The creation of a migrating path is necessary to enable epithelial cells to migrate through the fibrin clot or alongside the junction between the clot and underlying connective tissue, or the mucoperiosteal flap surface during periodontal surgery. Plasmin is an enzyme derived from the activation of plasminogen in the clot. Plasmin dissolves the fibrin barrier, thus facilitating the creation of the migration pathway. The migrating keratinocytes upregulate two activators: tissue-type plasminogen activator and urokinase-type plasminogen activator along with its receptor.9
Plasminogen exerts a significant role in wound re-epithelialization. This is emphasized by the failure of wounds in mice to re-epithelialize where the plasminogen gene has been knocked out. Plasmin is a fibrinolytic enzyme that leads to lysis of fibrin; the expression of plasmin and various other proteases facilitate cell migration by creating a path for the migrating cells.9
Matrix metalloproteinases (MMP) comprise a family of enzymes, and the function of each family member is to specifically cleave a subset of matrix proteins. The basal cells that have migrated past the free edge of the basil lamina secrete MMP-1, which is also referred to as interstitial collagenase. Native collagens are broken down by MMP-1, which facilitates cell migration by degrading Type I and III collagen.9
MMP-9, also called gelatinase B, cleaves type IV collagen in basal lamina as well as the type VII collagen that forms anchoring fibrils, and in so doing releases the basal cells from their adherence to the basal lamina. MMP-10 is also named stromelysin-2. It is proposed to have a wide spectrum of substrate specificity and it is expressed in wounds.9
The formation of granulation tissue is normally initiated approximately 4 days after wounding occurs. Primarily, it is composed of new capillaries, macrophages, fibroblasts and some loose connective tissue. Granulation tissue is a complicated repository of cytokines that demonstrate chemoattractive, mitogenic and other regulatory functions.9
Growth factors are also found in granulation tissue. They are derived mostly from macrophages at this stage of healing. Contingent upon the predominant cytokines and the stage of the granulation tissue, many activities occur such as cell proliferation, chemotaxis, and phenotypic expression of cells.9 It is important to remember that the final phenotype expressed by the cells is influenced to a large degree by the behavior of certain cell types and the nature of the extracellular matrix environment. As new granulation tissue is developing, macrophages, fibroblasts and new blood vessels grow into the wound space in a synchronized process. The fact that macrophages release cytokines that induce fibroblasts to synthesize an extracellular matrix attests to their interdependence. The extracellular matrix provides support for cell and vascular in-growth carrying nutrients to sustain the cellular functions.9
Fibroplasia is a term that refers to the component of granulation tissue comprised primarily of fibroblasts and the extracellular matrix they produce. Not only do macrophages produce cytokines, but fibroblasts also produce many cytokines and they respond to them in an autocrine mode. Cytokines stimulate fibroblasts to proliferate and modulate the production of extracellular matrix. Growth factors also participate in wound fibroplasia. The accelerated formation of granulation tissue is seen in wounds treated with purified growth factors.9
During wound healing, the extracellular matrix and fibroblasts interact in a reciprocal mode. The adhesion and migration of fibroblasts in the granulation tissue are promoted by elements of the extracellular matrix such as fibronectin and collagen. Fibroblasts produce and modulate the extracellular matrix, and the extracellular matrix regulates the expression of genes and the general behavior of fibroblasts. Integrin receptors enable fibroblasts to adhere to fibronectin, collagen and vitronectin.9
In preparation for migration, fibroblasts, like keratinocytes, have to rearrange their integrin expression profiles. In healthy tissue, fibroblasts are located in matrices that are abundant with collagen. Fibroblasts located in the vicinity of the wound respond to wounding by downregulating the integrin receptors for collagen and upregulating the integrin receptors necessary for adhesion to elements of the provisional matrix such as fibrin, fibronectin and vitronectin.9
Fibroblasts demonstrate an extraordinary ability to respond to signals from the extracellular environment. When fibroblasts are stimulated concurrently by signals from both provisional matrix (fibrin or fibronectin) and growth factors (such as platelet-derived growth factor) fibroblasts selectively upregulate the receptors for provisional matrix components. In contrast, when fibroblasts are challenged by platelet-derived growth factor in the presence of a collagenous matrix, the fibroblasts upregulate the receptors for collagen and but not the receptors for the provisional matrix.9
Little is known about how extracellular matrix molecules in the wound environment affect the regulation of fibroblast gene expression. Even more problematical is the fact that some extracellular matrix molecules appear as different isoforms as a result of alternative splicing of their transcripts at the pre-messenger RNA level.9 An isoform is a protein that has the same function and similar (or identical sequence), but it is the product of a different gene and is usually tissue specific.3
The familiar molecule fibronectin appears as different isoforms because the alternatively spliced domains, EIIIB and EIIIA, in its gene structure are spliced in or out. Whereas the fibronectin from plasma lacks both of the two spliced domains, the fibronectin produced by fibroblasts and macrophages in wounds includes the spliced domains EIIIB and EIIIA. Surprisingly, the isoform of fibronectin containing EIIIB and EIIIA that is found in healing wounds, is also the principal form of fibronectin in embryonic tissues where cell migration is occurring.9
Yet, it is not completely understood what function is fulfilled by including the spliced domains EIIIB and EIIIA in fibronectin during embryonic development and wound healing. More recent studies proposed that the insertion of these domains into the fibronectin molecule may promote the migration of cells by regulating the adhesiveness of the substrata. In addition the CS-III domain, also a spliced domain of fibronectin, may possibly facilitate the migration of cells in wounds.9
The integrin receptor α4β1 is utilized by fibroblasts to bind to the CS-III domain of fibronectin.Cell culture studies have demonstrated that that the expression of the integrin a4b1 facilitates migration, whereas movement can be impeded by the standard fibronectin receptor a5b1 that binds to the RGD sequence. Studies revealed that fibroblasts found in the healing wound may markedly over express the spliced domain CS-III.9
Fibroblasts demonstrate an amplified expression of the classic a5b1 integrin once cells have completed their migration into the wound. During migration, fibroblasts in wounds directly utilize vitronectin and fibrin as substrata for adhesion. The availability of cell membrane receptors for these matrix proteins enhances this process.9
Lastly, growth factors such as platelet-derived growth factor and transforming growth factor-b indirectly stimulate the migration of fibroblasts in wounds by upregulating some of the integrin receptors mentioned above.9 As wound healing continues, fibroblasts migrating into the wound synthesize a new, collagen-rich matrix which they use to replace the provisional matrix. Fibroblasts synthesize specific extracellular matrix molecules in the wound; this is regulated by transforming growth factor-b1 and some other growth factors, their sources, and effects listed in Table 19.
| Growth Factor | Source | Effect |
| Fibroblast growth factors 1, 2, 4 | Macrophages, endothelial cells | Fibroblast proliferation and angiogenesis |
| Transforming growth factor-a | Macrophages, keratinocytes | Re-epithelialization |
| Transforming growth factors b1, 2 | Platelets, macrophages | Fibroblast and macrophage chemotaxis; extracellular matrix synthesis; secretion of protease inhibitors |
| Epidermal growth factor | Platelets | Re-epithelialization |
| Platelet-derived growth factor (isoforms AA, AB and BB) | Platelets, macrophages, keratinocytes | Fibroblast and macrophage chemotaxis, fibroblast proliferation and matrix synthesis |
| Keratinocyte growth factor | Dermal fibroblasts | Keratinocyte proliferation |
| Insulin-like growth factor | Plasma, platelets | Endothelial and fibroblast proliferation |
| Vascular endothelial growth factor | Keratinocytes, macrophages | Angiogenesis |
| Interleukin 1alpha, beta | Neutrophils | Activate growth factor expression in macrophages, keratinocytes and fibroblasts |
| Tumor necrosis factor-alpha | Neutrophils | Activate growth factor expression in macrophages,
keratinocytes and fibroblasts |
The expression of collagenous matrices in wounds can also be induced by cytokines such as interleukin-4. As soon as the obligatory amount of collagenous matrix has been synthesized, signals are produced that stimulate down-regulation of collagen synthesis; however, these signals are not well understood. Some researchers have proposed that the collagen matrix itself may exert some type of regulation.
About 7–10 days after wounding has occurred, some of the fibroblasts found in the wound change into myofibroblasts. The myofibroblasts then express a-smooth muscle actin, producing strong contractile forces leading to wound contraction. Programmed cell death then decreases the number of fibroblasts and myofibroblasts in the healing wound during the final stages of fibroplasia.9
Embryonic wounds heal without a lot of contraction and scarring in contrast to wounds in adult tissues. Fibroblasts do not transform into myofibroblasts in embryos and the angiogenic response is diminished. Wounds in embryos demonstrate low but temporary levels of expression of transforming growth factor-ß1 compared with wounds in adult tissues.9
Transforming growth factor-ß1 has been associated with pathofibrotic conditions and this may account for the propensity of adult wounds to respond to the healing process with scarring and contraction. Antibodies have been shown to neutralize the effects of transforming growth factor-ß1 in healing wounds; and when these antibodies are applied to wounds, scarring has been diminished.9
The term angiogenesis refers to the formation of new blood vessels, especially new capillaries; it involves the process of vascularization of tissue; and it comprises an essential event during wound healing.3, 9 It is the red granular appearance of new blood vessels that invade the healing connective tissues in wounds that led to the conception of the term ‘granulation tissues’. As noted in Table 1, various growth factors play a significant role in stimulating blood vessel growth in healing wounds.9
Macrophages and damaged endothelial cells synthesize fibroblast growth factor-2, and wound-edge keratinocytes and macrophages induce vascular endothelial growth factor. Studies have been done where researchers have depleted fibroblast growth factor-2 with a consequent inhibition of angiogenesis in the wound.9
Similar to fibroblasts and keratinocytes, endothelial cells must upregulate certain integrins (aVb3) on their cellular surface to enable them to respond to angiogenic signals. Unfortunately, none of the signals that induce proliferation, migration and phenotype expression in endothelial cells are thoroughly understood as yet.9
Obviously angiogenesis is a complicated mechanism that depends on the availability of a suitable matrix in the wound. Endothelial cells first migrate into the provisional matrix; they then form tubes surrounded by their own provisional matrix; that is subsequently followed by a true basement membrane. In order to maintain a balanced process, like fibroblasts, endothelial cells associated with angiogenesis in wounds must also go through programmed cell death at the same time as the matrix matures; this is characterized by regression of capillaries.9
Therapy used to treat tissues damaged by periodontal disease usually results in healing by repair. If inherent tissue regeneration is to occur at wound margins, the elimination of causal factors, inflammation, and pockets is essential. Guided tissue regeneration warrants important consideration in the treatment plan for periodontal tissue regeneration due to its capacity to form a connective tissue attachment. Yet, there are limits to the amount of potentially newly formed connective tissue attachment and bone that can be regenerated because there is limited proliferation of the cells. It is the goal of research to promote the proliferation, migration, and differentiation of the cells that possess the ability to form new connective tissue attachment.8
Scientists have conducted detailed studies on the biological regulation of periodontal tissue regeneration in vitro and applied the findings in vivo. Studies have demonstrated that the proliferation, movement, attachment, and differentiation of cultured cells are regulated by a variety of molecules such as platelet-derived growth factor, insulin-like growth factor-I, basic fibroblast growth factor and fibronectin, or factors contained in cementum or dentin.8
It was assumed that these factors would have a useful clinical application in periodontal regenerative procedures. Indeed, some studies have demonstrated the legitimacy of applying factors to periodontal wounds, while others have disclaimed their effectiveness in periodontal regeneration. Normally, the molecules in factors are prepared in the wound, but molecules that are applied externally may be degraded immediately by their related enzymes. This problem may be solved by including hybrid molecules in the application technique.8
Root planing to debride the root surface in combination with acid etching removes the smear layer from the denuded root surface. This exposes type I collagen fibers; they are chemotactic to fibroblasts that produce collagen, ultimately connecting with the newly formed collagen fibers. Acid root conditioning was once believed to facilitate the attachment of connective tissue to the root surface. More recent studies have indicated that acid etching is not particularly effective in promoting the formation of new connective tissue. Further studies are needed on the techniques for periodontal tissue regeneration, such as guided tissue regeneration and fibronectin application in combination with acid etching.8
At the outset, use of autogenous cancellous bone and marrow in bone grafting was described as forming new bone and connective tissue attachment. Newer studies revealed that osseous grafting contributed minimal predictable regeneration of periodontal tissues. Yet some studies demonstrate its usefulness.8
In contrast to grafting with bone, more recent studies utilized hydroxyapatite, tricalcium phosphate, polylactic acid, collagen, etc., substances that are artificial fillers. Fillers containing growth or differentiation factors, for instance bone morphogenetic protein, have also used to graft bone defects. The complications of antigenicity and infection are resolved by the use of synthetic fillers. Unfortunately, the results from grafting with synthetic fillers have not been consistent.8
If the granulation tissue is unable to actively proliferate among the fillers, bone grafting and its substitutes will be precluded, since the newly regenerating epithelial cells will migrate on the surface of the granulation tissue. If the bone regenerates unexpectedly, whether conductively or inductively, and makes contact with the tooth surface, ankylosis results.8 “Osteoinduction is the process by which osteogenesis is induced. It is a phenomenon regularly seen in any type of bone healing process. Osteoinduction implies the recruitment of immature cells and the stimulation of these cells to develop into preosteoblasts. In a bone healing situation such as a fracture, the majority of bone healing is dependent on osteoinduction. Osteoconduction means that bone grows on a surface. This phenomenon is regularly seen in the case of bone implants. Implant materials of low biocompatibility such as copper, silver and bone cement shows little or no osteoconduction. Osseointegration is the stable anchorage of an implant achieved by direct bone-to-implant contact. In craniofacial implantology, this mode of anchorage is the only one for which high success rates have been reported. Ingrowth of bone in a porous-coated prosthesis may or may not represent osseointegration.”24
Researchers questioned whether bone grafting materials or artificial fillers were capable of stimulating the formation of new connective tissue attachment on the tooth surface. They recommended that research be undertaken to develop absorbable fillers capable of stimulating osteoinduction and maintaining enough space to promote selective proliferation of periodontal ligament-derived cells on the denuded root surface.8
To reiterate, periodontal ligament-derived cells retain their properties in vitro. Preliminary results indicated that cultured cells derived from the periodontal ligament were capable of producing new periodontal ligaments when they were reimplanted with a demineralized root in vitro. Utilizing fibroblast cells derived from a periodontal ligament to seed a planed root surface resulted in considerable attachment gain. However, additional studies are warranted.8
Guided tissue regeneration is a form of periodontal therapy that employs the use of an occlusive membrane to selectively guide cells derived from the periodontal ligament to populate the denuded root surface. Cells derived from the periodontal ligament have been described primarily in vitro. These cells possess some characteristics normally found in osteoblasts and have phenotypes that can differentiate into osteoblasts or cementoblasts.8 The phenotype is defined as the total characteristics seen in an organism resulting from the interaction between the genotype and the environment regardless of the actual genotype of the organism.3
Various growth factors have been isolated from dentin or cementum. There is evidence that cells derived from the periodontal ligament have the capacity to form mineralized nodules in vitro, and that growth factors or proteoglycans regulate the proliferation, migration, and differentiation of these cells.8
Over the last 20 years, there has been an immense increase in the knowledge base about growth factors, cell adhesion molecules and cytokines, with a significant improvement in the understanding of the cellular and molecular biology of wound healing. A number of new techniques are now being utilized to influence the wound healing process. Enhanced reepithelialization in animal model burn wounds has been reported after the application of epidermal growth factor and transforming growth factor-a. Likewise, keratinocyte growth factor applied to skin wounds has resulted in mitogenic effects on the healing epithelium. Attempts have been made in the past to topically apply fibronectin to promote periodontal wound healing because cell adhesion and migration are essential to wound healing.9
However, caution is warranted in the use of recombinant molecules as exogenous sources to enhance wound healing. In considering fibronectin, the plasma form of fibronectin, which lacks the spliced domains EIIIB and EIIIA, is already present in ample amounts in the provisional matrix of a healing wound. Addition of exogenous quantities of the plasma form of fibronectin has no biological rationale and an improved outcome is not a realistic expectation.9
Researches still do not know the biological functions of the spliced domains EIIIB, EIIIA and the CS-III segments. Scientists are currently evaluating their roles. If they prove to specifically enhance cell migration, then particular isoforms of cell adhesion molecules that permit weak cell adhesion while promoting migration of cells would be valuable.9
In normal, healthy subjects, abundant amounts of growth factors are found in the provisional matrix. Supplementation with the proper growth factor may be advantageous in systemic conditions such as diabetes that exhibit a reduced availability of various growth factors. Since some growth factors may have a short half-life, exogenous augmentation of wounds with the correct growth factors applied in the later stages of healing by a delayed release mechanism might be helpful in healing wounds found in healthy subjects as well. In addition, initial study results indicate that the utilization of recombinant growth factors to treat osseous defects during periodontal surgery has potential.9
Research in the future must concentrate on understanding the molecular mechanisms of differential gene expression in wound healing. Wound healing is achieved by the carefully orchestrated biological interactions of inflammatory cells, keratinocytes, fibroblasts and endothelial cells, all in a coordinated response to a complicated variety of signals.9
A large number of these processes have start and stop signals. Paramount is a thorough understanding of these signals and their consequences of differential gene expression in responder cells; when this is accomplished, hopefully we can look forward to therapeutic management of wounds that results in true regeneration of damaged tissues.9
Periodontal tissues are naturally endowed with a high proliferative activity. However, extensive tissue damage often results from periodontal diseases which destroy the tissue framework, and the consequence is healing in the form of repair. Thus, accomplishing the regeneration of damaged periodontal tissue is very difficult.8
Based on the known concepts of periodontal wound healing and the regeneration of periodontal tissues, it may be possible to achieve the goal of periodontal tissue treatments by the following:8